Woodpigeon’s ( Columba palumbus ) autumn 2018 Migration : a particular research on a single “ corridor fly-way ” crossing Central Italy , and focus on “ flocking” , “hunting pressure”, “ age “ .
Woodpigeon’s ( Columba palumbus ) autumn 2018 Migration : a particular research on a single “ corridor fly-way ” crossing Central Italy , and focus on “ flocking” , “hunting pressure”, “ age “ .
Preliminary draft/report (July 2019) : updates the Text reported in http://journal.ilcolombaccio.it/flocking-preliminary-report-on-the-autumn-migration-2018-in-italy/Enrico Cavina .
Club Italiano del Colombaccio
con la collaborazione di Cacciatori/Rilevatori
Coordinatori : Rinaldo Bucchi e Denis Bianchi ( vedi Bibliografia)
Baggioni Stefano , Barboni Alberto, Bianchi Renato , Bigi Fabio, Bombardi Massimo , Borghesi Roberto , Boscherini Luigi ,Bucci Maurizio , Bucchi Rossano , Buzzi Andrea, Ceccarelli Morris , Cecchini Renato , Cenni Paolo, Conca Franco , Cresti Luca , De Naia Sandro, Gessi Franco, Gori Franco , Guidi Matteo, Lampronti Federico , Lanzini Alessandro, Lorenzoni Angelo, Mazzei Flavio , Monesi Lorenzo , Panzini Renzo, Pasqui Serafino ,Picchi Silvestro , Palli Enrico Francesco, Porcù Paolo , Ristori Gianni , Romei Ivan,Rossi Giuseppe, Rossi Riccardo , Rossinelli Alessio, Sangiorgi Domenico,Troni Alessio, Valenti Mauro,Valentini Roberto .
Un ringraziamento particolare a Renato Bianchi,Maurizio Bucci , Renato Cecchini,Paolo Cenni ,Lorenzo Monesi,Riccardo Rossi che hanno voluto collaborare con contatti diretti .
Ci scusiamo per eventuali omissioni e/o inesattezze .
contact Email : ecavinaster@gmail.com
ABSTRACT ( english)
Many methods to study the migrations of birds have been developed along the ancient and modern history of the Ornithology and Ecological Sciences .
The ringing ,as historical yet used method, is the most famous , and more recently we find radioisotopic tracking,DNA examination,radar monitoring of migrating mass of birds ,direct following of some migrating birds by light aircrafts ,satellite radiotracking the most recent and promising, and we are in front of the evolution of the nano-thecnology inside the bodies of migrating birds . All these methods present limits of interpretation of the Migration’s phenomenon and behaviours , but their contribute is still basic in the modern knowledge of the birds’ Migration .
The most simple method to observe the migration is that when a man observes the migrating birds in the sky or in stop-over sites , but the limit is the visual field of observation : collecting data of local or extended observations was an evolution of so simple method ,and also it must be considered the base of all traditional migrations’ knowledge of hunters and more recently of organized birdwatchers , developing so important experiences also in the field of the so-called “CitizenScience”.
Collected data of all these methodologies were the base of other sophisticated methods of analysis by theorical ( statistics) and practical (laboratory and experiments) study’s evolution . Despite so impressive increasing of high qualified experiences , the birds’ Migration is still full of undiscovered misteries and questions without answers. Collecting so many data by records for many birds’ species and in many different timing and locations , often not well coordinated , it seems many times too far from the real behaviours ,choises and decision making of birds before,during,after the migrations. But visual observation,recording of numbers of birds and flocks, and their instant and long- term behaviours in front of habitat and climatologic changes , seems the most “live” method in real time .
The opportunity to collect and study data , recorded by very expert and traditional observers/hunters along a single narrow long (300 Km) migration’s corridor/route that crosses Italy by Woodpigeons as short-medium-long distance migrators ,seemed to us a new method powered by the solution of continuity following the migration along their full crossing way and autumn timing .
In the present study on a migratory corridor/route ( long 300 Km-large 15-25 Km ) the observations of recorded birds (woodpigeon- Columba palumbus ) were 496.389 in 3.975 flocks , but considering an enlarged area in Central Tuscany ( para-corridor) were 607.749 in 5.805 flocks ( 1st October-15 November 2018 ) . We have also considered detailed analysis during the two “waves/peaks” of the seasonal migration . 41 experts Observers has registered detailed data hour by hour , day by day . In the present preliminary draft/report we have consideterd 3 topics:
– Flocking : the sizes of flocks changes during the fly crossing Central Italy in the corridor “Mesola forest – Elba island” as by a sequence fom average values of 306 birds (Mesola) to 46 ( Central Tuscany) and then 81 and finally 156 ( increasing 239%) in Elba island .
– Hunting pressure : it results 0,52 % ( numbers of killed birds 3.201)
– Age : in the first period (1st Oct.-17 Oct)of the autumn migration 1358 YOUNG birds are 55,96% , and in the second period 1843 birds are 50,24% .
Many detailed comments are reported in Italian Text- please use Translators in your Language.
Please take note the the present preliminary report is simply a DRAFT
. The results of the present study draft/preliminary report wll be basis of future statistical study ( work in progress)
IL CORRIDOIO

IL CORRIDOIO in TOSCANA con aree PARA-CORRIDOIO

PUNTO DI ARRIVO sul DELTA del PO – MESOLA
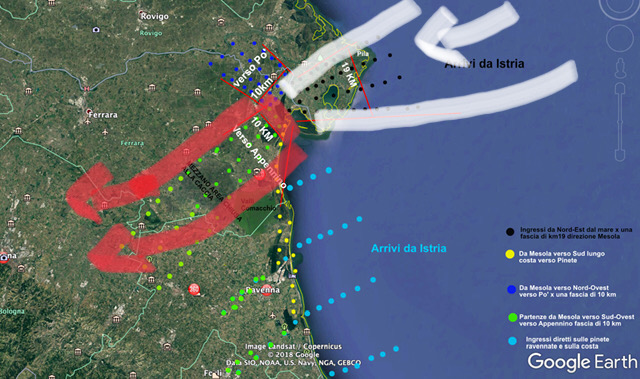
Come più volte segnalato – e non da ultimo sulla relazione-pubblicazione “Poggibonsi 2019 :Progetto Colombaccio Italia “ – Coordinatore Rinaldo Bucchi – sulla base di libretti e contributi diretti selezionati ( 41),è in stato di analisi avanzata tutto l’andamento della migrazione autunnale su un “Corridoio migratorio” ristretto ,molto ristretto ( di ampiezza 10-25 Km ) per una “direttrice” preferenziale che corre dal Delta del Po (Mesola) all’Isola d’Elba ,attraversando da Nord Est a Sud Ovest tutto il Centro Italia (Emilia-Romagna e Toscana ) .
I dati raccolti da Cacciatori/Rilevatori selezionati anche nell’ambito di PCI,MSM,MCL dovrebbero consentire ( work in progress) di analizzare dettagli della Migrazione riguardanti vari aspetti dell’eco-sistema migratorio del Colombaccio , e qui in anteprima riportiamo quelli riguardanti la composizione numerica degli stormi in migrazione ( flocking ) e successivamente l’analisi dell’effetto venatorio e dell’età dei Colombacci abbattuti ed esaminati direttamente dai Rilevatori.
.
Il metodo ( un singolo “corridoio”) mai applicato su grandi numeri ed in sequenza di monitoraggio stagionale nella Ricerca Ornitologica ,è innovativo ed intrigante : è come cercare di capire cosa fa e come si muove in un mese e mezzo ( 1 Ottobre-15 Novembre ) tutta la massa migrante ( composta da popolazioni ad origini diverse sconosciute e con varie ondate ) che sbarca in un punto preciso d’arrivo Adriatico ( Delta del Po-Mesola) e –su una Direttrice preferenziale- parte poi ad attraversare tutto il Centro Italia sino ad un ulteriore preciso punto di partenza (Isola d’Elba) per l’ulteriore traversata trans-Tirrenica-Mediterranea .
Non è questa la sede per descrivere e discutere i dettagli metodologici-scientifici di questa Ricerca “in corso” , ma alcuni dati già ora sono molto significativi e possono interessare molti appassionati oltre che gratificare i selezionati Osservatori-Rilevatori che hanno partecipato alla raccolta dei dati in analisi . Il lavoro di estrazione (attenta puntigliosa lettura dei Libretti dei Rilevatori ) ,archiviazione, sistematizzazione e schematizzazione di questi dati è stato ed è lungo, impegnativo e difficile ,ma alcuni risultati preliminari –per ora qui focalizzati sul “ flocking” – possono essere offerti alla discussione , magari essa stessa preliminare di Lavori a stesura scientifica più impegnativa .
Sono stati registrati –con buona se non ottima approssimazione – 496.389 Colombacci in 3975 voli di stormi ,tutti specificatamente osservati nel Corridoio geografico in esame da Mesola ad Elba ,tutti con lo stesso metodo di conteggio

Va ben considerato che il Corridoio Mesola-Elba (CME) è molto ristretto ( ampiezza 10-25 Km) e se rapportato a tutta la lunghezza peninsulare ( specie catena Appenninica) che accoglie il transito migratorio , si quantifica per valori minimi percentuali sul 2-3 % di tutta detta lunghezza da Nord a Sud : offre però il vantaggio di poter seguire la Migrazione con i dati registrati quasi “in diretta cronologica” ogni ora ed ogni giorno su una ben precisa “direttrice” preferenziale .
L’analisi si è sviluppata – e si svilupperà ulteriormente ( work in progress ) – su 5 sub-segmenti di registrazione inclusi in 2 segmenti base ( Est – Ovest ) :
2 sub-segmenti nell’area orientale pre-Appenninica ( Mesola – Emilia Romagna );
3 sub-segmenti nell’area occidentale post-Appenninica ( Mugello- Toscana centrale e costiera- isola d’Elba ).
Nel tragitto pre-Appenninico (2 sub-segmenti ) sono stati registrati 209.319 Colombacci in 1371 voli.
Nel tragitto post-Appenninico (3 sub-segmenti) sono stati registrati 287.319 Colombacci in 2604 voli.
MUGELLO
Nel tragitto pre-Appenninico e nel primo sub-segmento post-Appenninico ( area Mugello ) non vi era alcuna possibilità di duplicazione di conteggio .
Nel tragitto post-Appenninico ( Toscana centrale e costiera ) ,invece , esisteva questa possibilità di duplicazione , eccettuato il transito specifico sull’Isola d’Elba dove è stata effettuata ( Renato Cecchini ) una precisa “scrematura della duplicazione” escludendosi quindi duplicazioni di conteggio su tre direttrici in arrivo-transito-partenza sull’isola .
Rimanendo base del Lavoro il ristretto corridoio geografico Mesola-Elba (ampiezza 10-25 km),si è ritenuto opportuno considerare in “appendice” anche le aree Toscane “para-corridoio” , limitrofe cioè ad Ovest-Est del corridoio stesso in 11 siti selezionati di osservazione , risultando le medesime importanti aree di stop-over dopo l’attraversamento delle creste Appenniniche e relativa dispersione “a ventaglio” sulla Toscana , prima del ricompattarsi poi di tutta la “massa migrante” proiettata all’attraversamento del Mar Tirreno .
Deve essere chiaro che questo allargamento di conteggio di 11 siti “para-corridoio” –comunque considerato affetto da possibili duplicazioni di conteggio – si offre a critiche e speculazioni interpretative di possibili “variabili” , come ad es. da più numerosi arrivi rispetto alla “porta Mugello”, caratteristiche delle aree di stop-over, orografia locale, memoria geografica, meteorologia orografica ecc. . Ci è sembrato corretto questo “allargamento para-corridoio” ,statisticamente abusivo ma pur sempre complementare di una corretta “lettura” della Migrazione .
Questo “allargamento para-corridoio” porta ad un aumento dei Colombacci e voli registrati : rispettivamente 607.749 e 5.805.
La “lettura” della Migrazione globale su “corridoio” e “para-corridoio” ed ambedue insieme , si avvale anche delle analisi di dettaglio nelle 2 Ondate/Picchi identificati nella stagione 2018 ( 10-15 Ottobre // 20-22 Ottobre ) secondo i parametri espressi nel ns. precedente Lavoro basilare “ The general pattern of seasonal dynamics of the Autumn migration of the wood pigeon ( Columba palumbus)in Italy ” ( pubblicato in The Ring 40,10.1515-2018 ).
I dati Ondate/Picchi 2018 sul “corridoio” e “ corridoio + para-corridoio “ , per 5+3 = 8 gg. ( su 46 gg.del periodo- da escludere giornate di silenzio venatorio ) così si quantificano :
– 273.000 Colombacci in 2044 voli ( valori degli 8 giorni Ondate/Picchi che corrispondono rispettivamente al 58,53 % e 51,42 % di tutta la Migrazione in un mese e mezzo )nel “corridoio” esclusivo .
363.498 Colombacci in 3339 voli ( 59,80 % e 56,54 % di tutta la Migrazione ) in “ corridoio + para-corridoio” .
Nel complesso la percentuali di Colombacci migrati attraverso l’Italia Centrale indicano che quasi il 60% della massa migrante nell’autunno 2018 si è concentrata ( giorni registrati) negli otto giorni delle due Ondate/Picchi sul totale dei 46 giorni del periodo ( 1 Ottobre-15 Novembre ) . Purtroppo dobbiamo ancora una volta rilevare come la mancanza di dati certi circa le ORIGINI delle varie POPOLAZIONI ,che nidificano nel Paleartico Occidentale ed oltre la catena degli Urali (Russia,Scandinavia,Bielorussia,Europa Nord-Orientale, area Carpatica e Balcanica ), non permette di identificare la dinamica dei movimenti migratori da tali differenziate aree di origine . In ragione di questo deficit di base è impossibile analizzare specifici aspetti di Ecologia migratoria , quali dipendenza dall’evoluzione temporale climatica ed ambientale , dinamica del “decision making” del momento d’involo migratorio , scelta della fly-way migratoria principale , effetti del raggrupparsi di popolazioni diverse in aree di stop-over , varie condizioni meteo-atmosferiche determinanti gli spostamenti dalle aree di origine , modifiche permanenti o meno dei comportamenti migratori ( migratori a breve,media,lunga distanza . tendenza a divenire stanziali , inurbamento ) .Tutto ciò all’insegna del carattere principale della Specie e cioè la “flessibilità” comportamentale e tutto ciò potrà forse trovare risposte solo quando saranno svolte ricerche con Radioisotopi , studio dei DNA territoriali , collaterali indicazioni dal Radiotracking satellitare .
FLOCKING
Tutto quanto sinora esposto permette già ora di documentare ( e riteniamo non sia mai stato fatto ,così come risulta dalla ns. accurata ricerca in Letteratura) un Tema di Ricerca ( “ flocking” ) particolarmente interessante e tuttora poco approfondito , che è quello dell’entità numerica degli stormi in migrazione ,quale si modifica durante il percorso migratorio “ arrivo-transito-stop.over-partenza “ .
Questa fenomenologia di gruppo , che risulta importante aspetto di studio della c.d. Ecologia sensoriale e comportamentale delle Specie animali migratrici , ben si documenta nelle caratteristiche proprie migratorie del Colombaccio ( Columba palumbus ) durante l’attraversamento della nostra penisola ed i nostri dati vogliono rappresentarsi in termini di documentazione “evidence based” ineccepibile .
Fatta base numerica di “quanti Colombacci stimati in uno stormo “ in rapporto con “ quanti Colombacci in totale abbiamo osservato” otteniamo la/le MEDIE di entità del/dei branco/branchi in varie condizioni spaziali e temporali .
Nella nostra analisi preliminare su un corridoio ristretto ( spazio) nei giorni di migrazione (tempo ) di una singola stagione abbiamo potuto estrarre i seguenti dati :
Un valore ( “media” ) di 306,23 per stormo-Colombacci all’arrivo sulla costa Adriatica ( Mesola);
Un valore di 128,45 subito prima lo svalicamento dell’Appennino ,quindi un’iniziale frammentazione dei branchi ;
Un valore di 46,29 subito dopo lo svalicamento dell’Appennino ,quindi una ulteriore drammatica frammentazione dei branchi;
Un valore di 81,60 nella fase di dispersione (Toscana) di transito/stop-over sino alle coste,quindi un graduale ricompattarsi in branchi via via più grossi rimanendo nello spazio “corridoio”;
Un valore di 103,57 se aggiungiamo le aree “para-corridoio” in Toscana , quindi un effetto di ulteriore agglomerazione nelle aree di stop-over ,se pur breve ;
Un valore di 156,12 nella fase di transito/partenza dell’Isola d’Elba ,quindi un definitivo ricompattamento in stormi sempre più grandi protesi all’attraversamento del Mar Tirreno .
La sequenza delle “medie” (numero Colombacci per branco ) da Mesola ad Elba è quindi : 306,23 – 128,45 ( decremento del 58,2% )– 46,29 (decremento ulteriore Mugello del 64,1% ) – 81,60 ( incremento 76,1%) – 103,57 ( ulteriore incremento del 27,2 % ) – 156,12 ( dal minimo di 46,29 media in Mugello l’incremento è del 239,1 % )
Quantificando in percentuale i valori di “ decremento/incremento” sul percorso completo ricaviamo un decremento dell’85% da Mesola a Mugello , e successivamente un incremento del 239% alla partenza dall’Elba .



Per la corretta “lettura” di questi pochi ma sicuri dati sul “ flocking-deflocking- reflocking “ in un corridoio ristretto trans-peninsulare in Italia , in una stagione accuratamente monitorata da 41 Rilevatori , non possiamo escludere che necessiterà di un approfondimento critico in dipendenza delle “variabili” contingenti .
Comunque questa nostra indagine – qui presentata in anteprima – con un focus di osservazione molto ristretto ( il 2-3 % di tutto il tragitto trasversale peninsulare ) documenta ed indica chiaramente che i Colombacci arrivano con stormi più grandi dopo aver attraversato il Mar Adriatico ,si frammentano via via nel percorso orografico e durante brevi stop-over, tendono poi a ricompattarsi in branchi sempre più grossi per attraversare il Mar Tirreno e più largamente il Mediterraneo .
Particolarmente significativa per queste fluttuazioni di dimensione degli stormi ,è l’evidenziazione in dettaglio di quanto accaduto in alcune giornate 2018 di Ondate/Picchi quando l’intensità dello stimolo migratorio è massimo ( furor) .
Nella prima giornata di Ondata del 10 Ottobre ( 18.230) la media di dimensione dei branchi si rappresentava in questa sequenza:
area Mesola 385 uccelli ,area pre-Appenninica 79 , area post-Appenninica Mesola 30 , corridoio Toscano 57 , para-corridoio 44 , Elba 128 . ( EST-OVEST 385-79-30-57-44-128 ) ( primo decremento -92,2 % – ulteriore incremento 326,7 % )
Il 13 Ottobre (26.055 ) la sequenza è : 712-42 ( decremento -94,1% ) -38-46-60-98 ( incremento 157,9 % )
Il 14 Ottobre (41.933) : Mugello 37-Toscana Centr. 92-58 – Elba 121 ( incremento 227 %)
Il 15 Ottobre ( 63.165) : Corridoio Toscano 76 , para-Corridoio 31 – Elba 145 ( incremento 90,8%)
Nella seconda Ondata il 20 Ottobre ( 84.822) : Mugello 67 – centro Toscana 137-51 – Elba 223 ( incremento 232,8 % )
Si inserisce in questa documentazione il fenomeno delle “strisce” chilometriche osservate da noi in Italia ( vedi decolli di massa da Mesola,S.Rossore – segnalazioni Appenniniche )
ma anche a Falsterbo ( Svezia), Portland (Canale della Manica UK) , Pirenei (Francia-Spagna) specie nei percorsi in mare aperto . Ricordiamo che il fenomeno era ricorrente nelle migrazioni della Colomba viaggiatrice (Passenger) ora estinta in Nord America .
I motivi di questo comportamento “sociale” sono vari , si configurano per una sfida interpretativa , rimangono nel mistero e rientrano nella Etologia-Ecologia di vita associativa di una Specie aviaria anche durante la migrazione ,così come emerge da alcuni importanti non numerosi Lavori ai quali rimandiamo il Lettore in Bibliografia e Web-bibliografia .

Questa suesposta “anteprima” vuole solo essere un richiamo alla necessità di approfondire gli studi sulle ORIGINI dei Colombacci e connesse diverse popolazioni con probabili diversi comportamenti associativi prima,durante,dopo la Migrazione .
Lo studio interpretativo del “flocking” inteso come “dinamismo associativo animale” richiede –per chi è interessato ad approfondire – una NOTA aggiuntiva specifica alla quale rimandiamo ( vedi in calce con relativa Bibliografia ).
EFFETTO “ CACCIA
La Caccia in quanto tale ( predazione) incide negativamente sullo “status” o consevazione della Specie Columba palumbus in Italia ?
I dati raccolti in questa Ricerca indicano chiaramente “NO” ( incidenza 0,52%) e si allineano ad altri numerosi rilievi statistici riportati in Lavori pubblicati o citati in IJWR . Altrimenti per paradosso è possibile anche affermare che la Caccia incide troppo poco rispetto all’aumento generalizzato delle popolazioni del Paleartico Occidentale e oltre , nel contesto di modificazioni ambientali per lo più legate all’attuale fase di antropizzazione in molte aree europee . In una supposta proiezione di “secoli” dobbiamo in qualche modo ricordare che in un passato recente (1800-1900) in presenza di una crescita esponenziale nel Nord America la Columba migratoria (Passenger pigeon) si è estinta molto verosimilmente per un processo di auto-sterilizzazione della Specie ( danno-modifica DNA ? ) .
Affrontando nel merito l’effetto “caccia” nella particolare indagine/analisi condotta sulla massa migrante ( autunno 2018) di Colombacci ( Columba palumbus ) in un ristretto ( raggio 10-15 km) corridoio sulla direttrice Mesola-Elba con la registrazione ( 41 Rilevatori esperti selezionati ) di 496.389 Colombacci in 3975 voli , aumentata a 607.749 in 5850 voli sul corridoio allargato ad aree di stop-over dopo lo svalicamento dell’Appennino ,oltre al fattore “ dimensione numerica “ degli stormi (flocking ) ed oltre altri fattori in futura analisi , abbiamo analizzato in dettaglio e qui riportiamo :
– numero dei Colombacci abbattuti e raccolti , confrontati con il numero globale di quelli osservati e registrati ( effetto Caccia ) ;
– età differenziata ( stimata con parametri univoci come da precise indicazioni reperibili nel collegato MSM 2018 vedi https://www.ilcolombaccio.it/CMS/wp-content/uploads/2018/02/MSM-completo.pdf ).
DA MSM( Rinaldo Bucchi) testo e libretti : oltre la valutazione del collare ( assenza nei giovani , incompleto negli immaturi – questi quindi ambedue colombacci di annata – completo negli adulti )
Nella parte esterna delle ali hanno risalto le penne “remiganti” che sono 10 e per semplicità numerate da 1 a 10 (la penna remigante più esterna è contrassegnata dal numero 10). Ogni numero corrisponde alla progressività della muta delle penne (prima mutano le remiganti interne e l’ultima a mutare sarà pertanto la n. 10). Le remiganti sono ricoperte superiormente dalle cosiddette “copritrici alari” ed anche queste piume mutano dall’interno all’esterno. Per distinguere un colombaccio giovane da un adulto occorrerà verificare il colore delle copritrici alari che se saranno grigiastre e bordate di marrone apparterranno ad un giovane, se piuttosto nerastre saranno di un adulto. Quindi: a copritrici alari grigie bordate di marrone corrisponde un soggetto giovane. Per distinguere un adulto da un giovane ci sono altri parametri da verificare: i giovani hanno zampe color marrone, gli adulti color rossiccio. Di seguito: il becco dei giovani è cartilagineo e di color marrone, quello degli adulti coriaceo di color avorio – giallastro – a volte anche rosato. Alla stessa maniera l’iride dell’occhio dei vecchi è gialla, quella dei giovani marrone. Anche il peso dei soggetti esaminati può suggerire l’età degli stessi: i vecchi sono molto più pesanti dei giovani/giovanissimi. Infine le macchie bianche iniziano ad ornare il collo dei colombacci all’età di 4/5/6 mesi e sono molto appariscenti dopo l’anno d’età dei soggetti presi in esame.
La base è rappresentata da 3.201 Colombacci .
L’attività venatoria incide quindi nello 0,52 % della migrazione globale di 607.749 Colombacci osservati / registrati .
Questi dati si allineano perfettamente con la tendenza globale della Caccia registrata in altri lavori riportati e citati in IJWR , per più anni e per grandi numeri che viene sempre a collocarsi in una forbice 0,50-2 % .
La “lettura” dell’incidenza della caccia sulla migrazione , al di là di schemi statistici, si avvale anche del controllo cronologico dei Libretti dei Rilevatori , dove l’andamento delle attività venatorie emerge anche in termini discorsivi .
I dati attuali 2018 sono anche indirattamente indicativi del fatto che l’incidenza della caccia aumenta modicamente nelle aree con stop-over dopo lo svalicamento dell’Appennino nella prima fase di migrazione anziché nella seconda fase ( 17 Ottobre-15 Novembre) quando più forte è la spinta migratoria , così come le percentuali di abbattimento si riducono drasticamente nel punto d’inizio (isola d’Elba) della transvolata del Mar Tirreno e Mediterraneo , quando gli stormi si sono di nuovo ingrossati ed il “furor” migratorio è al massimo
Altro elemento appunto da considerare è la differenziazione tra 2 periodi ( 1-17 Ottobre / 18 Ottobre-15 Novembre ) e 2 fasi di Ondate/Picchi , in particolare anche per la valutazione “età”.
Nel primo periodo ( 1-17 Ottobre) comprensivo dell’Ondata/Picco ( 10-15 Ottobre) sono stati abbattuti nel “ corridoio + para-corridoio “ 1358 Colombacci e di questi 699 ( 51%) nel corridoio e 659 (49%) nel para-corridoio.
Nel secondo periodo ( 18 Ottobre-15 Novembre ) globalmente 1843 Colombacci e di questi 1201 (65 %) nel corridoio e 642 (35%) nel para-corridoio ( Toscana centrale)
Il tutto su un totale di 3201 Colombacci abbattuti e raccolti .
ETA’
Analizzando i dati globali l’ulteriore indagine permette di raccogliere dati precisi –anche nella fase di raccolta registrazione e valutazione dei sigoli Rilevatori – e dati molto indicativi sull’andamento della migrazione dei Colombacci nel rapporto “ GIOVANI / ADULTI “ . La valutazione dell’età come precedentemente esposto ha principalmente catalogato nei Libretti tre categorie “giovanissimi,giovani immaturi ,adulti” : ai fini della esclusiva analisi migratoria abbiamo ritenuto di considerare prioritarie due sole categorie , basandoci sul timing della nidificazione ed esperienza o meno di una precedente Migrazione “ Giovani/Adulti” .
I dati raccolti ci permettono la seguente schematizzazione :
– nel primo periodo ( 1 Ottobre-17 Ottobre) 375 GIOVANI nel corridoio ristretto e 385 nel para.corridoio per un titale di 760 GIOVANI ( 23,74% di 3201 totale);
– nel secondo periodo (18 Ottobre-15 Novembre) 575 GIOVANI in corridoio e 349 in paracorridoio per un totale di 926 GIOVANI ( 28,92% di 3201 totale)
Quanto sopra per quanto riguarda i Giovani mentre per gli Adulti :
– nel primo periodo 324 ADULTI in corridoio e 274 in para-corridoio per un totale di 598 ADULTI ( 18,68% di 3201 totale);
– nel secondo periodo 626 ADULTI in corridoio e 293 in para-corridoio per un totale di 917 ADULTI ( 28,64% di 3201 totale )
Nel primo periodo abbiamo 1358 ( 42,42% di 3201) dove i GIOVANI sono il 55,96%.
Nel secondo periodo abbiamo 1843 ( 57,57% di 3201) dove i GIOVANI sono il 50,24%.
Dobbiamo sottolineare e ricordare che il numero “3201” rappresenta solo lo 0,52% di 607.749 Colombacci registrati in volo . Nella valutazione sia pur minimale dato il numero “piccolo” i GIOVANI sono apparentemente più numerosi – 55.96%-nel primo periodo della Migrazione e comunque ben presenti anche nel secondo periodo 50,24%.
Questi dati possono essere confrontati con alcuni dati ricavati da analoga Ricerca 2018 simultanea e condotta su 5 Direttrici differenziate con 22 Rilevatori ed un totale di 326.000 Colombacci registrati e 2.206 Colombacci abbattuti ( 0,67%) dove l’incidenza dei Giovani è di 1147 su 2206 cioè il 51,99% in confronto con 1059 Adulti .(MSM – Rinaldo Bucchi) https://www.ilcolombaccio.it/CMS/wp-content/uploads/2018/02/MSM-completo.pdf
Altri dati di dettaglio analitico – incrociati per tempo ( primo e secondo periodo della Migrazione autunnale ) e spazio ( corridoio ristretto – corridoio allargato – ambedue ) e relative numerose “variabili” – potrebbero essere riportati , ma di fatto non sembra possono offrire particolari chiavi di lettura della Migrazione , data anche la già citata minimale consistenza del “campione” (3201 = 0,52% della massa migrante ) ; ciò vale soprattutto per l’analisi cronologica e sequenziale dei movimenti di popolazioni a migrazione breve-media-lunga e relative fasi di nidificazione a monte del corridoio in Italia . Manca una base statistica valida sulla reale identificazione -anche geografica- di queste popolazioni .
Questo aspetto di mancata conoscenza delle precise ORIGINI delle popolazioni che arrivano da noi in diverse ondate come è stato evidenziato già con i dati di Progetto Colombaccio ( 1997-2007 – dieci anni ) ed esposto con approfondita analisi scientifica in “ The general pattern of seasonal dynamics of the Autumn migration of the wood pigeon ( Columba palumbus)in Italy ” ( pubblicato in The Ring 40,10.1515-2018 ) ci suggerisce alcune domande :

– quando arrivano da noi le popolazioni subito a ridosso degli Urali ( versante Asiatico e versante Europeo ) ? quando sono partite ? quali soste in aree di stop-over hanno fatto ?
– e questo vale per le popolazioni nidificanti in Russia,Bielorussia,Scandinavia,Ucraina , regione Carpatica ( Romania,Polonia,Slovacchia) ed ancora Ungheria, Nord e Centro Nord della penisola Balcanica e via via anche da Regioni più vicine a noi ?
– quali condizioni ambientali hanno influito sulla durata o meno di stop-over ? quali condizioni atmosferiche hanno determinato le partenze ?
– quali condizioni di muta e quali prevalenze di età si sono verificate nella tempistica di percorso ?
– quali –forse- sub-Specie possiamo ipotizzare migranti insieme alla massa migrante ( roscioli,topacchi,codoni )
– quali cariche di parassiti ( e potenzialmente anche virus ) possono realizzarsi e dove , durante il percorso ?
– il mischiarsi di queste popolazioni in varie aree di stop-over realizza movimenti continuativamente “mischiati” oppure le popolazioni con varie Origini mantengono una loro individualità di percorso prima,durante,dopo l’attraversamento dell’Italia ?
Tutto ciò rientra nel grande capitolo della Ecologia della Migrazione ed Ecologia sensitiva della Specie .
Queste ed anche altre domande stigmatizzano la necessità di una grande –quanto possibile – precisa Ricerca sulle ORIGINI .
CONCLUSIONI
In definitiva sulla base di quanto esposto possiamo forse azzardare – sempre nei termini di una Anteprima-Bozza preliminare – una qualche conclusione di questa particolare circoscritta analisi e focus su una Direttrice tradizionale / corridoio MESOLA – ELBA .
Flocking
I branchi o stormi giungono in Italia con grandi numeri per i branchi che attraversano il Mar Adriatico , poi si riducano gradualmente durante l’attraversamento peninsulare per poi ricompattarsi in grandi branchi al momento ripartire per attraversare il Mar Mediterraneo
Effetto “ Caccia “
L’incidenza di prelievo rimane costante tra 0,50-1 % .
Età
Pur con qualche variazione temporale nel nostro “campione” specifico la distribuzione tra Giovani e Adulti si mantiene equilibrata tra 50-55 % nelle varie focalizzazioni statistiche e nelle varie fasi della Migrazione . Ciò può significare che questo equilibrio è ulteriore conferma dell’ottimo stato della Specie Columba palumbus .
Ancora una volta dobbiamo sottolineare quanto sia grave la mancanza di uno studio specifico sullr Origini delle varie popolazioni che giungono in Italia secondo i parametri già identificati nel lavoro
“ The general pattern of seasonal dynamics of the Autumn migration of the wood pigeon ( Columba palumbus)in Italy ” ( pubblicato in The Ring 40,10.1515-2018 )
Questo quindi si rappresenta come una sfida importante della Ricerca nei prossimi anni , così come rimane non intrapresa la sfida della Ricerca di base anatomo-fisiologica sull’Organo Para-Timpanico di Vitali che è alla base delle conoscenze biologiche eco-sensitive .
Il nostro attuale Lavoro molto circoscritto e qui rappresentato con analisi preliminare , rimane aperto all’approfondimento specialistico “statistico” ( elaborazione ) che speriamo di poter svolgere con le opportune collaborazioni .
La Bibliografia rimane quella iscritta in
“ The general pattern of seasonal dynamics of the Autumn migration of the wood pigeon ( Columba palumbus)in Italy ” ( pubblicato in The Ring 40,10.1515-2018)
aggiornata alle Bibliografie iscritte nei Lavori in Italian Journal Woodpigeon Research
e più recentemente a :
Rinaldo Bucchi : Monitoraggio Selettivo Migrazione ( MSM) https://www.ilcolombaccio.it/CMS/wp-content/uploads/2018/02/MSM-completo.pdf
Denis Bianchi : RISERVA NATURALE BOSCO DELLA MESOLA : la migrazione, lo stop.over in ottobre e lo svernamento del colombaccio Anno 2018/2019. https://www.ilcolombaccio.it/CMS/prima-giornata-di-monitoraggio-svernanti-eseguita-il-11-dicembre-2018-dalla-squadra-romagnola-pochissimi-i-colombacci-avvistati-lungo-la-strada-statale-romea-nel-percorso-di-andata-verso-mesola-e-sola/
* * *
Ogni suggerimento,critica,commento di esperienza ,e quant’altro saranno particolarmente graditi.
E’ possibile che nella trascrizione ed elaborazioni di così tanti numeri siano stati commessi errori comunque marginali nel contesto analitico globale .
Enrico Cavina
ecavinaster@gmail.com
scritto nel luglio 2019
NOTA Aggiuntiva a :
“Flocking” : una Ricerca particolare sulla Migrazione 2018 –
Anteprima dei risultati su oltre mezzo milione di Colombacci (Columba palumbus) monitorati in un ristretto “corridoio” migratorio da Mesola ad isola d’Elba .
“ FLOCKING” ovvero “riunirsi in volo di gruppo” ovvero “ dinamiche degli stormi “
“ Flocking” è un termine in lingua Inglese che include “FL…” ( derivazione da “ Fly” ) e che definisce la tendenza di Uccelli a riunirsi in stormo ( gruppo,branco; flock in Inglese ) anche se poi a prescindere dalla collocazione aviaria viene utilizzato a volte abusivamente un po’ per tutto il mondo animale (dai batteri all’uomo ) per esprimere la tendenza a raggrupparsi , imbrancarsi ,riunirsi in gruppo e ciò vale nel mondo aviario sia per Uccelli migratori diurni o notturni sia per Uccelli sedentari .
I vari stimoli che determinano e gestiscono le dinamiche del “flocking” sono biologici ( metabolici, ormonali,genetici ) , fisici ( nel rapporto con l’ambiente esterno ) e comunque riferibili a più complesse interazioni di ecologia sensitiva e soggetti a dinamiche matematiche e fisiche come più recenti lavori di studio (ad esempio per i grandi voli ed evoluzioni degli storni ) hanno evidenziato ed analizzato .
Alcuni concetti interpretativi di base sono ben riportati in un Lavoro antico del 1952 : Emlen J.junior – Flocking behaviour in birds – Auk 69,160-170,1952 . Secondo questo Autore l’associarsi in gruppo si realizza come il risultato di una mutua attrazione tra individui singoli . La tendenza degli Uccelli a rispondere positivamente alla presenza di individui della stessa Specie ,ma a volte anche di Specie diverse, viene definita come “gregarismo” ,ma questo termine sarebbe troppo generico come esplicativo di varie dinamiche :infatti dovremmo considerare la “tolleranza” di rapporti in comune con altri individui ,che può essere anche “attrazione” –opportunista,strategica,di necessità – in contrasto con l’eventuale “intolleranza” ( repulsione,respingimento,antagonismo ) a condividere con altri il proprio spazio e le proprie attività come ad esempio il pasturare insieme per mangiare su un limitato territorio . Il “flocking” dovrebbe quindi considerarsi risultato complesso e dinamico di due forze ,una positiva una negativa .Il bilanciamento che si realizza tra queste due forze ,anche su basi genetiche o istintive , guiderebbe l’associazionismo di gruppo nel determinare diverse dimensioni e numero di individui per gruppo nel proprio spazio-ambiente , e nel dinamismo temporale contingente : ciò con differenze sostanziali che si evidenziano anche nelle caratteristiche comportamentali ad esempio tra uccelli sedentari e migratori . L’analisi di Elmen J. Si rifà ad una più ampia ed indefinita filosofia interpretativa della vita sociale animale , ma non manca di riferirsi ad elementi fisiologici , come influenze endocrine connesse anche alla stagionalità sessuale e riproduttiva nonché dipendendo da fattori grossolanamente fisici come lunghezza della luce diurna e risposta alle condizioni atmosferiche .
A seguire il Lavoro di Elmen J. altri ,ma non molti Ricercatori si sono occupati del “flocking” ,ma non senpre in un’analisi globale biologica ,quindi spesso focalizzandosi su aspetti –pur importanti – tearici atteinenti il proprio campo d’interesse scientifico come ad esempio matematica,fisica,aerodinamica . Nel 1987 Reynolds C.W. ha pubblicato “ Flocks,Herds and Schools : a distribuited Behaviour model” ( SIGRAPH ’87,Conference Proceedings pgg. 25-34,1987 definendo tre aspetti fondamentali del “flocking” :
accentramento di soggetti animali come desiderio di stare insieme e vicini ad altri animali ;
evitare le collisioni tra di loro ;
acquisire velocità e direzione di volo dei soggetti accanto.
Ulteriori più recenti studi ( Lee e Co-Auth. : Dynamics of prey-flock escaping behaviours in response to predators attack – Journal Theoret.Biology 2040,250-259,2006 ) hanno evidenziato come il comportamento di volo di gruppo è fortemente relazionato alla “difesa dai predatori” .Più recentemente (2012) Blomkvist O. in una Tesi d’Ingegneria Fisica ( Royal Insitute Technology –KTH Vetenskap Och Konst ) “ Mathematical modeling of flocking behaviour “ svolge un’analisi esaustiva in termini matematici focalizzandosi appunto su un insieme di forze che in varia alternanza determinano scenari di diverse comportamenti e quindi di strategie per difendersi da attacchi di predatori , e quindi “l’unione fa la forza”. Il tutto comandato da uno stato di attrazione-repulsione ( respingimento ) animale tra soggetti di singola Specie o di Specie diverse ( …val la pena di ricordare “ allodole e civetta”) .
Di certo da questi studi emerge la complessità del fenomeno “flocking” e le difficoltà ad interpretarlo compiutamente .
Sempre in termini di analisi ed interpretazioni teorico-pratiche ,un recentissimo (2018) Lavoro è stato prodotto in Italia : Cavagna A. ed al. –The physics of flocking : correlation as compass from experiments to theory – Physics Reports :728,1-62,2018 ( CNR – Roma ed Univ.La Sapienza Roma ) . Questo Lavoro – nel suo complesso non accessibile alla comprensione dei non-addetti-ai-lavori ( come noi) ,ma diremmo esclusivo per fisici,matematici,statistici – si avvale della osservazione dei movimenti spettacolari degli Storni e specifiche analisi di come si muovono gli immensi branchi in volo e quali forze ( fisico-fisiologiche) determinano la trasmissione dei comandi “reattivi” e la realizzazione simultanea di movimenti così complessi appunto nella simultaneità dei movimenti di gruppo . L’analisi –matematica e statistica- approfondisce la dinamica della trasmissione dell’informazione –comando ( ….quali inter-connessioni tra nervi,scambi elettrochimici ,centri nervosi centrali e periferici ,muscoli si realizzano in infinitesimali frazioni di secondo nei componenti la “massa” – “flock” in volo ?? ) tra individui dello stesso branco e conseguente movimento di gruppo spazio-temporale.
Possiamo forse noi trasferire questa analisi della dinamica di “gruppo in volo” anche improvvisa e spettacolare , riferendoci alle conoscenze che ci derivano dalle tradizioni venatorie . Le storiche tradizioni di caccia al Colombaccio con le reti ( Pirenei ,Cava dei Tirreni ) basate sull’osservazione del comportamento del branco di fronte al possibile attacco “da sotto” del predatore ( Sparviero o similare ) : il branco con sorprendente simultaneità si butta a capofitto a raschiare il terreno nel tentativo di non dare spazio al predatore “da sotto” . Personalmente ho assistito a questo fenomeno nel corso di caccia di valico . Nel caso di questa trasmissione di conoscenza tra osservazione in Natura e creazione di un sofisticato metodo di caccia , i Cacciatori alzano le reti ad hoc ed il branco ci finisce dentro . Il comportamento del branco –in questo caso- si rifà al fattore negativo di Emlen J. e cioè “ respingimento” del predatore in termini di strategia di volo . L’altro effetto – fattore positivo “attrazione” è invece alla base di tutte le tecniche di caccia tradizionale in Italia con i messaggi “ forza scendete qui che c’è da mangiare “ , “ forza scendete qui che si può riposare e star sicuri “ . Se i messaggi “attrattivi” sono mal eseguiti si trasformano in“repulsione”.
Indipendentemente dall’ “attrazione” il sistema può non funzionare se la forza in atto dell’istinto migratorio è molto grande ( furor) prevale il “respingimento” .
Molto dell’ “attrazione” dipende dall’insito segnale di disponibilità di cibo , e nella Caccie al campo ( svernamento,stop-over ) questo aspetto è fondamentale . Le modalità di “flocking” per utilizzo alimentare sono state ampiamente studiate da R.K. Murton ( 1965-1980 ) sui Colombacci residenti nelle Isole Britanniche ,anche in termini di gregarismo e di strategie di Specie per la salvaguardia di Specie e correlata evoluzione di Specie.
Altri aspetti del flocking riguardano i comportamenti al momento dell’ “appollo serale “ quando la necessità di ambiente sicuro per trascorrere la notte ,è prevalente . Ciò vale anche per i luoghi di abbeveramento .
In conclusione , possiamo riassumere alcuni fattori “evidence based” che motivano e determinano le varie fasi dinamiche e interattive del “flocking” , differenziate anche nelle varie condizioni migratorie .
Molti occhi e “riflessi” in gruppo , meglio e più presto possono identificare il pericolo di predatori
Il gruppo (flock) con il suo comportamento di risposta e/o di prevenzione può disorientare il predatore e metterlo fuori gioco . Comportamenti analoghi si riscontrano anche nei Pesci ( ad es. sardine) e nei Mammiferi ( ad esempio Bue muschiato).
Nel gruppo “lo stormo” multiple esperienze di più popolazioni,famiglie,singoli individui ( i vecchi e le gerarchie di gruppo ) possono meglio guidare ed orientare il viaggio migratorio sia in termini direzionali ( orientamento , riconoscimento e memoria geografica ) sia in termini temporali ( decisioni di decollo-partenze , stop-over temporanei , valutazione delle condizioni meteo ) sia in termini di dimensione e composizione del gruppo ( distribuzione occupazionale delle aree di stop-over e disponibilità alimentare , guida del gruppo in volo , scelta dell’altezza di volo ).
Le risposte fisiologiche ,risposte a varie condizioni ambientali ed atmosferiche (PTO), di molti individui tutti insieme , si coagulano in una risposta di massa come decolli e transiti di massa .
Più esperienze tutte insieme possono facilitare erratismi alimentari in varie fasi di svernamento ..
Il gruppo nel suo insieme può meglio decidere il comportamento diurno o notturno in dipendenza della luce ( diurna o notturna “lunare”) e dello schermo “celeste-atmosferico” all’orizzonte ( nuvolosità,foschia,capacità di orientarsi con il sole e stelle )
Il gruppo nel suo insieme può meglio valutare (PTO altimetro ) la scelta di corridoi aerei di scorrimento migratorio e contingente valutazione del vento e dell’aerodinamica di volo anche in dipendenza dei sensori “meccano-recettori” delle penne ( più penne in più individui ovvero potenziamento di gruppo del sistema sensoriale ) e conseguenti scelte di “formazione” ( voli a “V”,palla compatta,striscia)
Il gruppo nel suo insieme può meglio esercitarsi nella previsione del tempo più o meno perturbato.
La continuità di comportamenti di gruppo può incidere sulle potenzialità evolutive di Specie ,come nella trasformazione da volatori a lunga migrazione a volatori a corta migrazione ,od evoluzione in sedentarietà
.
Le condizioni di vita di gruppo numeroso si offrono all’interscambio genetico in termini di miglioramento della Specie.
Quanto emerge dall’analisi della Letteratura e dall’elencazione conoscitiva di vari fattori che sono alla base del “flocking” , elencazione pur incompleta per varie condizioni di dettagli comportamentali distinti tra varie Specie aviarie , ben definisce l’attualità di studio e di Ricerca sul “flocking” –in particolare quello migratorio – pur anche riferito alla Specie Colombaccio ( Columba palumbus ).
Molti,molti elementi delle molteplici condizioni di stimoli vari che determinano caratteristiche e modalità di comportamento spaziale e temporale , rimangono sconosciute , ancor più tra i misteri delle Migrazioni degli Uccelli .
La miglior conoscenza del “flocking” rimane sfida alla Ricerca biologica,fisiologica,ecologica delle Specie aviarie .
Anche i nostri “evidence based” risultati derivati da una piccola Ricerca sulla migrazione selezionata 2018 del Colombaccio ( Columba palumbus ) , segnalano che le dimostrate modificazioni di entità di composizione degli stormi lungo il percorso Mesola-Elba si prestano ad ulteriori approfondimenti di studio .
Perché i Colombacci arrivano in grossi branchi dopo aver attraversato il Mar Adriatico , poi dopo stop-over o meno si frammentano via via lungo il percorso trans-peninsulare per poi ricompattarsi in branchi più grandi al momento di attraversare verso Ovest – Sud Ovest il Mar Tirreno ed il Mediterraneo verso i territori di svernamento ?
Quali popolazioni e con quali Origini tuttora sconosciute partecipano a queste formazioni di gruppo e con quali modalità di mischiarsi di singoli soggetti e/o famiglie e di diverse età ?
Molti misteri rimangono da esplorare .
PS – volendo divagare sul comportamento degli Uccelli in termini di territorialità e di flocking “aggressivo” , possiamo ricordare un Film “horror” di Hitchoch “UCCELLI” dove l’aggressività di gruppo viene come sublimata con una geniale sceneggiatura che rimane ingabbiata nella impossibilità di dare risposte adeguate del perchè questa aggressività e flocking su territorio , lasciando lo spettatore nella sua libertà d’interpretare come vuole , ma in definitiva abbandonandolo tra i misteri della Natura aviaria ed umana .
BIBLIOGRAFIA – WEB-BIBLIOGRAFIA ( oltre alle citazioni ne Testi)
https://www.sciencedirect.com/science/article/pii/S000334720900296
https://www.britishecologicalsociety.org/100papers/100_Ecological_Papers/100_Influential_Papers_008.pdf
https://www.sciencedirect.com/science/article/pii/S0003347214002747
https://www.sciencedirect.com/science/article/pii/S0022519302930651
https://www.sciencedirect.com/science/article/pii/S0960982215012932
https://www.sciencedirect.com/science/article/pii/S0003347209002966
https://www.researchgate.net/requests/attachment/55450562. *****
The physics of flocking: Correlation as a compass from experiments to theory
Andrea Cavagna a,*, Irene Giardina b,a,c, Tomás S. Grigera d,e,f
a Istituto Sistemi Complessi, Consiglio Nazionale delle Ricerche, 00185 Rome, Italy
A. Cavagna et al. / Physics Reports 728 (2018) 1–62
article info
Article history:
Accepted 16 November 2017 Available online 8 December 2017 Editor: Massimo Vergassola
abstract
Collective behavior in biological systems is a complex topic, to say the least. It runs wildly across scales in both space and time, involving taxonomically vastly different organisms, from bacteria and cell clusters, to insect swarms and up to vertebrate groups. It entails concepts as diverse as coordination, emergence, interaction, information, cooperation, decision-making, and synchronization. Amid this jumble, however, we cannot help noting many similarities between collective behavior in biological systems and collective behavior in statistical physics, even though none of these organisms remotely looks like an Ising spin. Such similarities, though somewhat qualitative, are startling, and regard mostly the emergence of global dynamical patterns qualitatively different from individual behavior, and the development of system-level order from local interactions. It is therefore tempting to describe collective behavior in biology within the conceptual framework of statistical physics, in the hope to extend to this new fascinating field at least part of the great predictive power of theoretical physics. In this review we propose that the conceptual cornerstone of this ambitious program be that of correlation. To illustrate this idea we address the case of collective behavior in bird flocks. Two key threads emerge, as two sides of one single story: the presence of scale-free correlations and the dynamical mechanism of information transfer. We discuss first static correlations in starling flocks, in particular the experimental finding of their scale-free nature, the formulation of models that account for this fact using maximum entropy, and the relation of scale-free correlations to information transfer. This is followed by a dynamic treatment of information propagation (propagation of turns across a flock), starting with a discussion of experimental results and following with possible theoretical explanations of those, which require the addition of behavioral inertia to existing theories of flocking. We finish with the definition and analysis of space– time correlations and their relevance to the detection of inertial behavior in the absence of external perturbations.
https://www.sciencedirect.com/science/article/pii/S0003347209002966
https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1474-919X.1974.tb07649.x
altre voci bibliografiche :
“Employing the Flocking Behavior of Birds for Controlling :Congestion in Autonomous Decentralized Networks” di Pavlos Antoniou, Student Member, IEEE, Andreas Pitsillides, Senior Member, IEEE,Tim Blackwell, Member, IEEE, and Andries Engelbrecht, Senior Member, IEEE
“Evolutionary Divergence in Brain Size between Migratory and Resident Birds”di Daniel Sol* e coll.
TUTTI I CACCIATORI / RILEVATORI che sono di diritto Co-AUTORI del Lavoro di RICERCA
“ “Flocking” : una Ricerca particolare sulla Migrazione 2018 – Anteprima dei risultati su oltre mezzo milione di Colombacci (Columba palumbus) monitorati in un ristretto “corridoio” migratorio da Mesola ad isola d’Elba .”
Coordinatori : Rinaldo Bucchi e Denis Bianchi
Baggioni Stefano , Barboni Alberto, Bianchi Renato , Bigi Fabio, Bombardi Massimo , Borghesi Roberto , Boscherini Luigi ,Bucci Maurizio , Bucchi Rossano , Buzzi Andrea, Ceccarelli Morris , Cecchini Renato , Cenni Paolo, Conca Franco , Cresti Luca , De Naia Sandro, Gessi Franco, Gori Franco , Guidi Matteo, Lampronti Federico , Lanzini Alessandro, Lorenzoni Angelo, Mazzei Flavio , Monesi Lorenzo , Panzini Renzo, Pasqui Serafino ,Picchi Silvestro , Palli Enrico Francesco, Porcù Paolo , Ristori Gianni , Romei Ivan,Rossi Giuseppe, Rossi Riccardo , Rossinelli Alessio, Sangiorgi Domenico,Troni Alessio, Valenti Mauro,Valentini Roberto .
Ci scusiamo per eventuali omissioni e/o inesattezze .
Un ringraziamento particolare a Renato Bianchi,Massimo Bucci , Renato Cecchini,Paolo Cenni ,Lorenzo Monesi,Riccardo Rossi che hanno voluto collaborare con contatti diretti .